Trans-Synaptic Signaling Complexes Formed by Adhesion Molecules
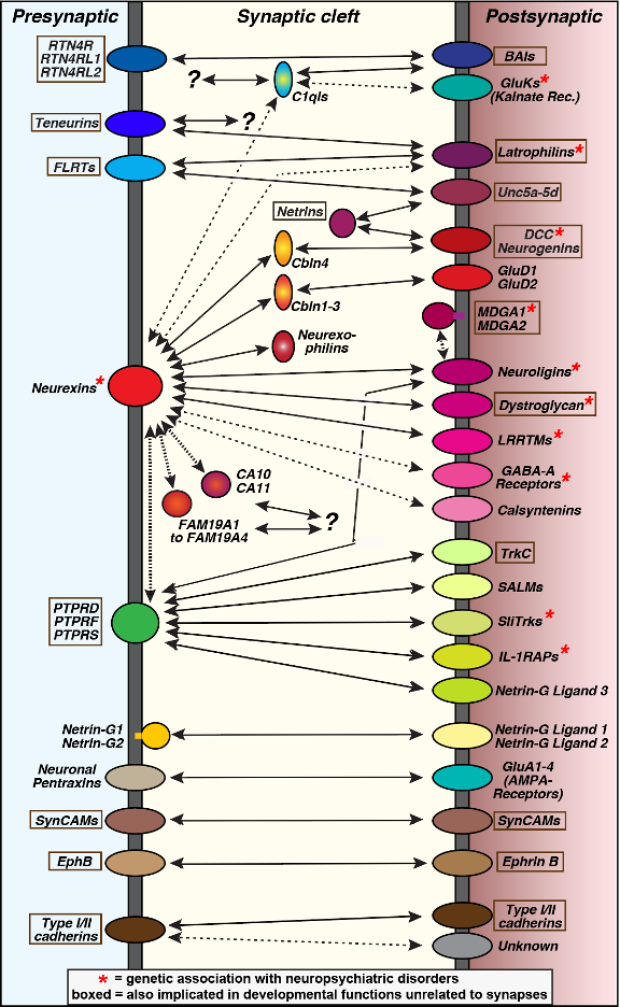
This page will provide a brief overview of trans-synaptic signaling complexes studied by the Südhof lab. As the field advanced and a multitude of trans-synaptic adhesion molecules were described, the need for a resource that summarizes the basic information on these complexes that emerged from the Südhof lab became apparent. The following information is neither exhaustive nor detailed but meant to provide a quick reference to the domain structures, salient functions and key open questions of a subset of synaptic adhesion complexes. The sections are organized according to key molecules such as neurexins or latrophilins, but this organization does not intend to imply that these key molecules are necessarily more important than some of their ligands, such as neuroligins and teneurins, or than other synaptic adhesion molecules.
The sections are organized according to three key molecules, neurexins, latrophilins, and Bai’s. This organization does imply that these molecules are more important than some of their ligands, such as neuroligins and teneurins, or than other synaptic adhesion molecules, such as LAR-type receptor phosphotyrosine phosphatases (PTPRD, PTPRF, and PTPRS) that are also presynaptic ‘hub’ molecules similar to neurexins.
As is apparent from this resource, the study of synaptic signaling complexes is only beginning. Although much is known, much more is unknown. Significant additional work is needed in order to understand the functional molecular architecture of synapses. At this point, a few common themes emerged that may help guide our thinking.
Core components vs. specifiers. A few families of synaptic adhesion molecules, such as neurexins, latrophilins, teneurins, neuroligins, and LAR-type PTPRs, appear to be invariably expressed in all neurons and may be present in all synapses, albeit at vastly different concentrations and in distinct variants, suggesting that they represent core synaptic adhesion molecules. Other families of synaptic adhesion molecules, such as neurexophilins, cerebellins, and DCC/neogenin, appear to be present only in a subset of neurons and synapses, suggesting that their actions mediate and specify those of the core components.
Interaction Networks. Neurexins, latrophilins, and Bai’s are each expressed from three genes whose products are diversified by alternative splicing and interact with multiple ligands. Many of these interactions can occur simultaneously, several of the interacting proteins are constitutive dimers or multimers, and occasionally interactions cross over between these three protein families. As a result, these adhesion molecules could form vast interaction networks in the synaptic cleft. Moreover, given the isoform-dependent differences in binding affinities and the activity-dependent regulation of the expression of some proteins and/or their splice variants, these interaction networks are likely dynamic and not static. These considerations suggest that trans-synaptic complexes form fluid interaction networks that can continuously change and will only be predictable once the precise concentrations of various components and their binding affinities are known.
Nanodomain clusters. Synaptic junctions are known to include a nanodomain substructure that organizes the positions of presynaptic vesicle docking sites and postsynaptic receptors2. Super-resolution microscopy revealed that synaptic adhesion molecules, as far as they have been studied, are also organized in nanoclusters, as shown for neurexin-1 and -3, teneurin-3, neuroligin-1 and -3, neurexin-3, LRRTM1, latrophilin-1, and Cbln13-8. Hippocampal synapses usually contain a single nanocluster but can have two or three clusters that generally occupy approximately 20% of a synaptic junction. Nanodomain clusters may represent signaling hubs in a synapse that organize the junctions.
Module & Motifs. Synaptic adhesion molecules are composed of multiple autonomously folded interaction domains that can share properties but differ in their specifics. It is notable that on the one hand, different ligands binding to the same domains generally share no sequence homology and operate by distinct structural mechanisms even for the same binding sites, but that on the other hand a few motifs are shared between synaptic adhesion complexes, such as the C1q-like domains of cerebellins and C1ql’s and the heparan sulfate modifications of neurexins and glypicans.
Control by Alternative Splicing. Most synaptic adhesion molecules (with notable exceptions, e.g., cerebellins, C1ql’s) are subject to alternative splicing. Alternative splicing has been most extensively characterized for neurexins that are expressed in thousands of variants (reviewed in ref. 9). Strikingly, some of these sites of alternative splicing control the functions of their resident proteins. For example, neurexin alternative splicing at SS2 and at SS4 determines what ligands they bind to9, whereas the C-terminal latrophilin-3 alternative splicing dictates their G-protein and postsynaptic scaffold interactions10.
References
1. Südhof, T.C. (2021) The cell biology of synapse formation. J. of Cell Biology 220, e202103052.
2. Biederer T, Kaeser PS, Blanpied TA. Transcellular Nanoalignment of Synaptic Function. Neuron. 2017 Nov 1;96(3):680-696.
3. Trotter, J.H., Hao, J., Maxeiner, S., Tsetsenis, T., Liu, Z., Zhuang, X., and Südhof ,T.C. (2019) Synaptic Neurexin-1 Assembles into Dynamically Regulated Active Zone Nanoclusters. J. Cell Biology 218, 2677-2698.
4. Zhang, X., Lin, P.Y., Liakath-Ali, K, and Südhof, T.C. (2022) Teneurins Assemble into Presynaptic Nanoclusters that Promote Synapse Formation via Postsynaptic Non-Teneurin Ligands. Nature Comm. 13, 2297.
5. Lloyd BA, Han Y, Roth R, Zhang B, Aoto J. Neurexin-3 subsynaptic densities are spatially distinct from Neurexin-1 and essential for excitatory synapse nanoscale organization in the hippocampus. Nat Commun. 2023 Aug 5;14(1):4706.
6. Han, Y., Cao, R., Qin, L., Chen, L.Y., Tang, A.H., Südhof, T.C., and Zhang, B. (2022) Neuroligin-3 confines AMPA-receptors into nanoclusters, thereby controlling synaptic strength at the calyx of Held synapses. Science Advances, 17, eabo4173.
7. Nozawa K, Sogabe T, Hayashi A, Motohashi J, Miura E, Arai I, Yuzaki M. In vivo nanoscopic landscape of neurexin ligands underlying anterograde synapse specification. Neuron. 2022 Oct 5;110(19):3168-3185.
8. Matúš, D., Lopez, J.M., Sando, R.C., and Südhof, T.C. The Essential Role of Latrophilin-1 Adhesion GPCR Nanoclusters in Inhibitory Synapses (2023) BioRX Preprint DOI: https://doi.org/10.1101/2023.10.08.561368
9. Südhof, T.C. (2017) Synaptic Neurexin Complexes: A Molecular Code for the Logic of Neural Circuits. Cell 171, 745-769.
10. Wang, S., DeLeon, C., Sun, W., Quake, S.R., Roth, B.L., and Südhof, T.C. (2023) Alternative Splicing of Latrophilin-3 Controls Synapse Formation. Nature, in press.