Current Research
Our lab aims to understand the mechanisms that create and shape calcium signals as well as their consequences for cell behavior. We are particularly focused on store-operated calcium channels because of their critical functions and their highly unusual properties and mechanism of activation. We study these channels at the level of single molecules to single cells with an integrated array of techniques that includes patch-clamp electrophysiology; protein biochemistry, mutagenesis, and engineering; and multiple microscopies including live-cell calcium imaging, total internal reflection fluorescence (TIRF) and confocal microscopy, whole-cell and single-molecule FRET, and electron microscopy.
Molecular choreography of STIM and Orai
The prototypic store-operated channel is the Ca2+ release-activated Ca2+ (CRAC) channel. The CRAC channel is highly unusual in that it is assembled reversibly and "on demand" in response to Ca2+ release from the ER. STIM1, the ER calcium sensor, and Orai1, the pore-forming subunit of the CRAC channel, are freely mobile in resting cells when the ER calcium store is full. After a stimulus causes the release of calcium from the ER, STIM1 oligomerizes and becomes trapped at ER-plasma membrane contact sites ("junctions") by interacting with plasma membrane inositol phospholipids, and Orai1 is trapped by binding to STIM1. We study these processes directly using single-particle tracking of fluorescently labeled proteins. We are interested in how ER-PM junctions form and move within the cell and how accessory proteins that bind to STIM and Orai modulate the assembly of STIM and Orai complexes at these sites.
Selected references: Wu et al, 2006; Luik et al, 2006; Luik et al, 2008; Park et al, 2009; Hoover and Lewis, 2011; Wu et al, 2014

Unique functional properties of the CRAC channel
CRAC channels display a unique set of biophysical and pharmacological properties that distinguish it from all other channel types. The channel is exquisitely selective for calcium yet has an extremely low conductance rate (>100-fold lower than other calcium channels), and its activity is shaped in several critical ways by calcium (fast and slow inactivation, potentiation). We recently found that STIM1 interacts functionally with several residues in the channel pore to influence fast calcium-dependent inactivation, and that all six Orai1 subunits must bind STIM1 to produce the channel's high ion selectivity and low conductance. As more information about STIM1 and Orai1 structure becomes available, we are using Orai1 concatemers, mutagenesis, and patch-clamp techniques to understand the molecular mechanisms that create the CRAC channel's biophysical signature.
Selected references: Mullins et al, 2016a; Mullins et al, 2016b

Simultaneous TIRF microscopy and patch-clamp recording from a T cell
A nanoscale view of CRAC channel activation: conformational dynamics of STIM and Orai
Calcium store depletion causes STIM1 to accumulate at ER-PM junctions, where its "CRAC activation domain" (CAD) binds to Orai1 to trap and open the CRAC channel. However, little is known about the nanoscale events underlying this process. How is STIM1 folded in the resting state to prevent premature activation, and how does store depletion drive the conformational changes that enable the CAD structure to engage Orai? Recently we have shown that the CRAC channel functions as a hexamer of Orai1 subunits, and that all six subunits must bind STIM1 to open the channel. These new results raise important questions about how STIM1 engages six binding sites on the channel. What is the stoichiometry and structural basis of the STIM-Orai interaction? We are applying single-molecule FRET and biochemical techniques to define the conformational dynamics of STIM as it becomes activated and to identify the binding interface between STIM1 and Orai1.
Selected references: Park et al, 2009; Rana et al, 2015; Yen et al, 2016
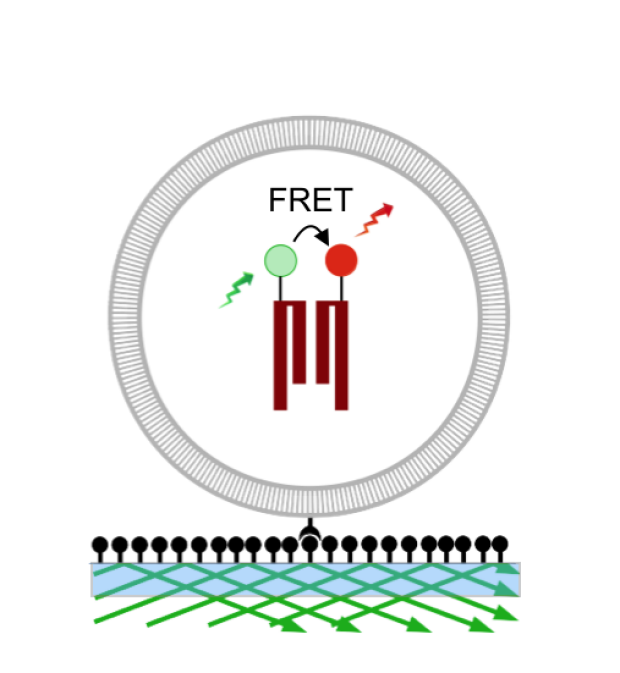

Single-molecule FRET of a labeled STIM1 cytosolic domain encapsulated in a liposome
CRAC channels and cell function
An adaptive immune response is initiated when a T lymphocyte forms a close contact -- an immunological synapse -- with an antigen-presenting cell. Engagement of the antigen receptor elicits a characteristic reorganization of the T cell's actin cytoskeleton at the synapse along with a dramatic flow of actin filaments from the periphery towards the center of the contact zone. These events are crucial for regulating the activity of the antigen receptor as it triggers downstream responses culminating in cell division and differentiation. We recently made a surprising discovery that calcium influx through CRAC channels is required for stimulating the reorganization and dynamics of actin at the synapse, and that the actin flow acts to concentrate the ER and CRAC channels in the center of the synapse, where exocytosis of cytokines and endocytosis of antigen receptors occurs. These effects on the cytoskeleton reveal a new target of calcium that is likely to modulate T cell activation in parallel with its well known action to regulate gene expression. We aim to understand the molecular underpinnings of these events and their specific impact on the cell biology of the immunological synapse.
Selected references: Hartzell et al, 2016
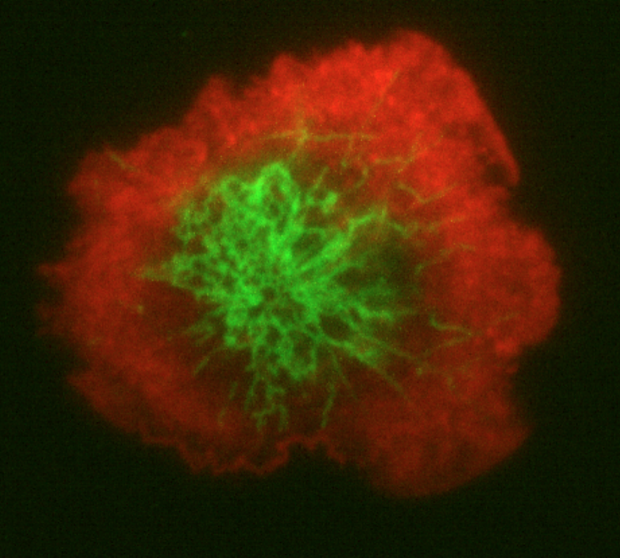
Organization of the ER (green) and actin (red) at the T cell synapse